



树突状(DC)是什么细胞,树突状细胞疫苗,DC生物细胞免疫疗法
树突状(DC)是什么细胞,树突状细胞疫苗,DC生物细胞免疫疗法
什么是树突细胞
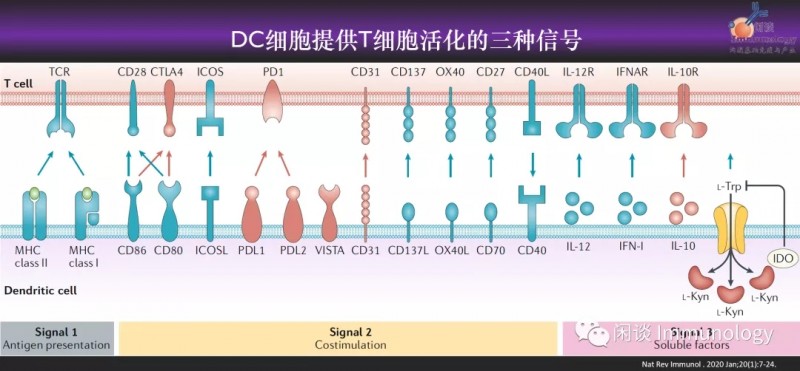
树突状细胞(dendritic cell,DC)作为专业的APC,提供给T细胞活化的第一,第二,第三信号。携带肿瘤抗原的DC疫苗,曾经给免疫治疗领域,带来很大的希望。但是临床上的响应率只有15%。
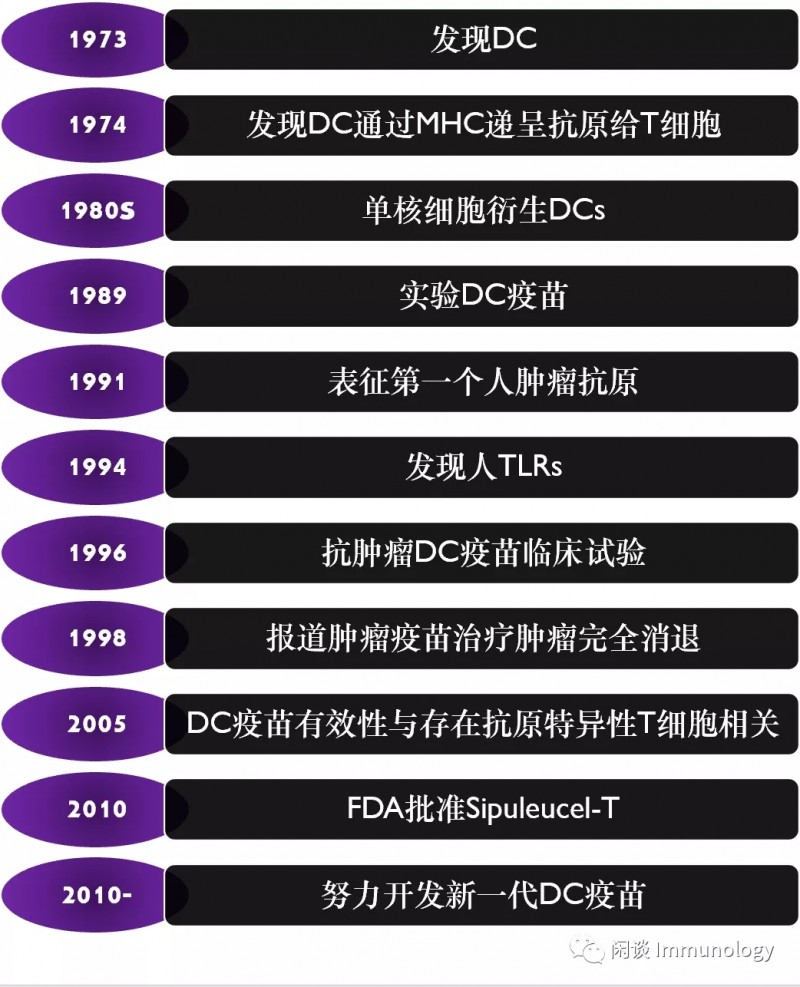
DC细胞研究简史
树突细胞的来源
树突细胞(DCs)是源自造血干细胞的异质性白细胞群体,其分化依赖于细胞因子Flt3L。DCs是一种特殊的抗原递呈细胞(APCs)群体,它们捕获抗原,处理并递呈给效应细胞,启动免疫反应。它们能够辅助初始T细胞和B细胞并以抗原依赖的方式调节免疫二次反应。DCs广泛存在于所有组织中,如骨髓、血液、脾脏、胸腺、扁桃体和淋巴结等二级淋巴器官。直到1973年Ralph Steinman和Zan Cohn的开创性研究才确定DCs是启动免疫反应所需的主要细胞亚群[1]。
树突细胞的亚型分类
DCs大致分为稳态、炎症性树突(Inflammatory DCs)和类似Langerhans细胞的亚群(图1)。稳态DCs可分为传统树突细胞(cDCs)和浆状树突细胞(pDCs)。cDCs又分为传统Ⅰ型树突细胞(cDC1)和传统Ⅱ型树突细胞(cDC2)。接下来小编将逐一为大家介绍DCs家族中每一位成员及其免疫作用。
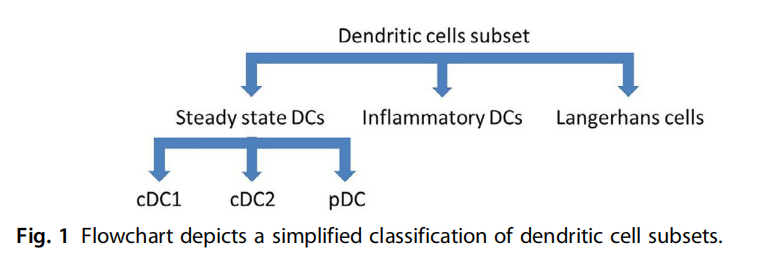
图1 树突状细胞亚群的简化分类
cDC1:cDC1具有独特的C型凝集素受体CLEC9A和趋化因子受体XCR1的表达特征。CLEC9A促进坏死细胞的吸收,有助于cDC1在生理条件下吸收死亡细胞,并将抗原交叉递呈给CD8 T细胞[2]。但在扁桃体和皮肤的cDC1上CLEC9A表达水平非常低。XCR1代表cDC1的未成熟期,在外周血和皮肤中可被观察到。cDC1通常被称为交叉递呈DCs,因其具有交叉递呈抗原的能力,并能引起抗肿瘤免疫反应[3]。DCs在人和鼠物种中具有保守模式,图2为人和鼠的cDC1表达marker基因示意图。
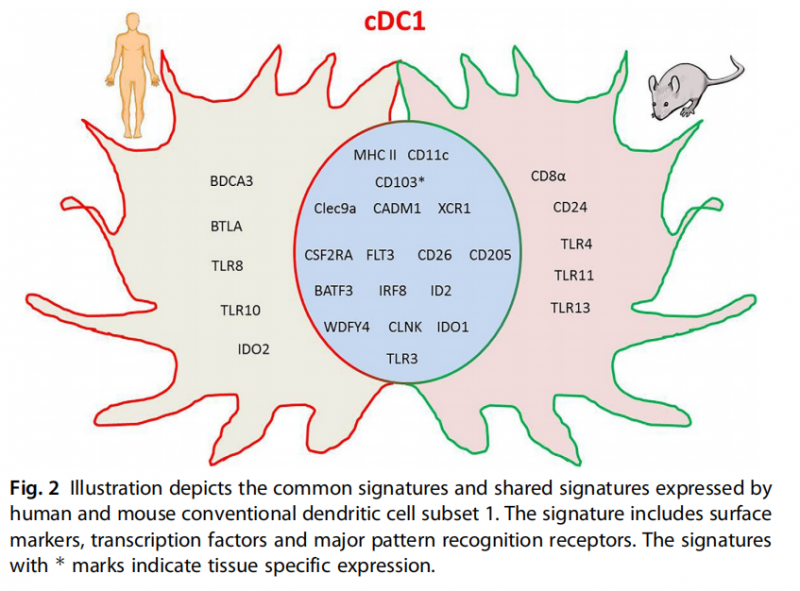
图2 人类和小鼠传统Ⅰ型树突状细胞所表达的共同marker。其特征包括表面标记物、转录因子和主要的模式识别受体。
cDC2:cDC2是通过高水平表达MHC Ⅱ、CD1C和SIRPA来识别的。肠内cDC2亚群表达更高水平的CD103。肺和皮肤表达CD1A,用CD64和CD14可区分皮肤单核吞噬细胞亚群与cDC2[4]。cDC2是存在于不同人体组织和器官中的主要DCs亚型群体。它们表达一系列转录因子如TLR,能够对从核苷酸到多糖的各种危险信号作出反应。与其他稳态DC亚群相比,cDC2还表达高水平的NLRPs和其他炎症相关信号分子,表明了感知不同危险信号时的特异性功能。图3为人和鼠的cDC2表达标志物示意图。
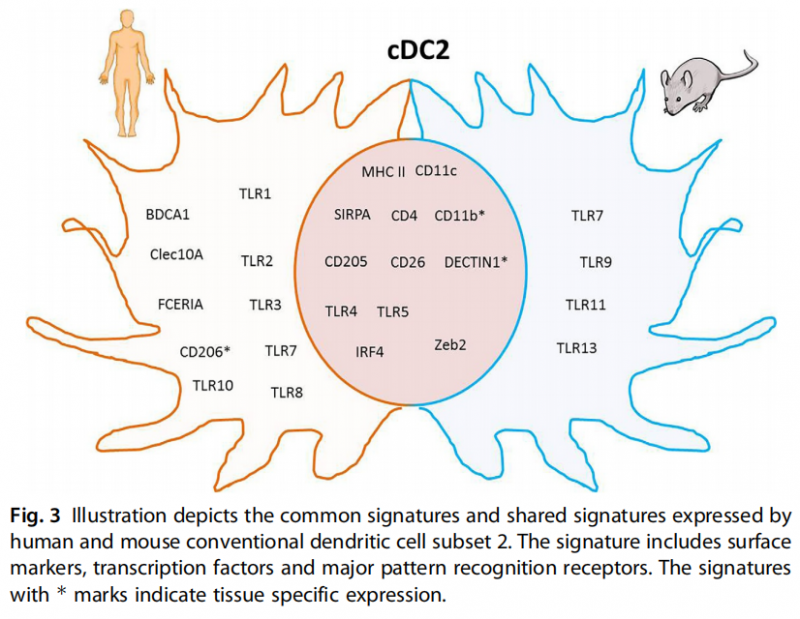
图3 描述了人类和小鼠传统Ⅱ型树突细胞所表达的共同标签。其特征包括表面标记物、转录因子和主要的模式识别受体。
pDCs:它具有独特的分泌细胞形态,如抗体分泌浆细胞,在稳定状态下缺乏典型的隐蔽的DC形态。pDCs是稳态DCs的第三种亚型,其独特的能力是产生数量极高的Ⅰ型干扰素(IFN),并在抗病毒免疫中发挥作用[5]。人类的pDCs的表达标记物包括CLEC4C、BDCA2、LILRA4和BTLA。小鼠的pDCs关键标记物是骨髓基质细胞抗原2(BST2)和唾液酸结合Ig样凝集素H(SiglecH)。在外周血中表达额外的标记物CCR9、SCA1和LY49Q。遇到病毒感染细胞时,pDCs是产生干扰素的细胞亚群,它们能够分泌Ⅰ型IFN和激活效应细胞的能力可能是抗病毒免疫的关键[6]。图4为人和鼠的pDC表达标志物示意图。
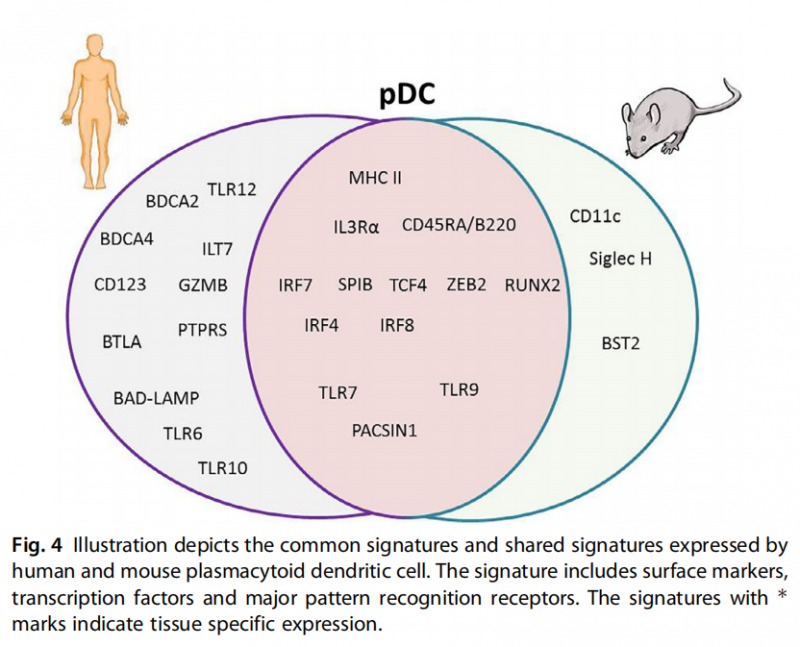
图4 描绘了人类和小鼠浆状树突细胞表达的共同标签。其特征包括表面标记物、转录因子和主要的模式识别受体。
Langerhans cells(LCs)
也称朗格汉斯细胞,LCs特异性分布于表皮组织,在所有物种中存在保守性,甚至在脊椎动物、鸟类和爬行动物中也有报道。因LCs不仅具有迁移到淋巴结的典型DC样特征,还能够递呈抗原并激活T细胞,所以被认为是DCs[7]。LCs具有典型的隐蔽形态,其中一个特征是存在Birbeck颗粒和表达Langerin或CD207。人类LCs表达人类白细胞抗原-DR(HLA-DR)、 CD1A和CD207。小鼠中表达CD24, CD11b和CD205,并且不表达CD26。LCs位于组织环境界面的最外层,是第一道免疫防御的一部分。LCs具有典型的隐蔽形态和长树突,常用于探测突破表皮层的角质层抗原。激活的LCs能够通过紧密连接向DCs扩展并获得外源抗原[8]。在稳态和炎症状态下迁移到淋巴结,并能启动免疫反应。
Inflammatory DCs
炎性树突细胞起源于单核细胞前体,在感染或损伤后,细胞因子或趋化因子产生,导致单核细胞前体分化,获得DC表型[9]。它们通常与巨噬细胞难以区分,表达高水平的MHCⅡ和不同的共刺激分子和CD11C。与其他DC亚群一样,它们可以在炎症反应时出现在任何组织中,并能够迁移到引流淋巴结。它们表现出CCR2依赖性迁移到淋巴组织,也能够向T细胞递呈抗原并引起免疫反应。
在免疫治疗中树突细胞的作用
DCs亚群在大多数健康和恶性组织中都有发现。乳腺癌、卵巢癌、肺癌、黑素瘤和肝细胞癌中均存在DC亚群。树突细胞存在于肿瘤微环境(TME)中,但是存在抗原交叉递呈受损,导致免疫无反应性的问题。缺氧、腺苷积累、乳酸水平升高和pH值降低均可损害DCs的正常功能。其中,脂质在DCs内的异常积聚是导致DCs功能障碍的主要机制之一。pDCs倾向于耐受性,有利于肿瘤进展,往往伴有不良预后。最近有研究表明,来自肿瘤引流淋巴结(dLNs)的CD103+DCs比来自非dLNs的DCs表达更高的PD-L1,阻断PD-L1和PD-1可减轻DCs功能障碍,从而引起TNFα、IL-12、IL-1β的产生和增加,增强了DCs刺激T细胞的能力[10]。在乳腺癌患者的单核细胞衍生的DCs对T淋巴细胞增殖的刺激作用较差,但对调节性T细胞(Tregs)的诱导作用较好,对DC功能评估可成为监测疾病进展的工具。
单细胞测序揭示树突细胞的作用机理
在2020年有研究报道称在慢性阻塞性肺病(COPD)患者的肺中传统树突细胞(DCs)能够协调淋巴样新生[11]。DCs在三级淋巴器官(TLO)形成和维持中的作用非常重要。然而,DC如何诱导TLO,尤其是在慢性阻塞性肺疾病(COPD)期间通过单细胞RNA测序研究表明人肺DCs诱导Tfh样细胞极化从而促进随后的淋巴新生。并证明了cDC2是诱导IL-21+ Tfh样细胞最有效的亚群。cDC2表现出一种独特的迁移特征,能够迁移到TLO形成的位置,并随后与CD4+T细胞相互作用。从COPD肺中分离的cDC2显示出更高的诱导Tfh样细胞极化的潜能。
DC疫苗研发要素
肿瘤抗原
荷载肿瘤抗原是DC疫苗的第一步。
分类:
●肿瘤特异性抗原(TSA):突变基因编码/ 新抗原,病毒基因,或癌症生殖系基因
●肿瘤相关抗原(TAAs):组织特异性基因编码或在肿瘤细胞中过度表达(和正常细胞相比)
来源:
●合成抗原肽
●抗原编码RNA/DNA
●自体或异体全瘤细胞裂解物
●肿瘤源全RNA/DNA
●肿瘤-DC杂交
挑战
●抗原在肿瘤表达的异质性
●递呈抗原肽与T细胞TCR亲和力低
DC成熟的Cocktails
多种促进DC细胞成熟的Cocktails在实验中使用如:
●促炎症因子(TNF,IL-1β,IL-6,PEG2等)+TLRs激动剂
●TNF+ IL1b + IFNg + IFNa + poly(I:C)
●电转染活化TLR4,CD40L,CD70
肿瘤负荷和肿瘤微环境
肿瘤释放抑制性细胞因子,MHC I类分子下调,肿瘤微环境中抑,TAMs,MDSCs,预示着DC疫苗效果不好。
手术,放化疗,靶向治疗降低肿瘤负荷,可增强DC疫苗效果。
DC疫苗针对小残留病灶效果好于原发或者转移病灶。
注射路径
●多靠近表浅淋巴结皮内注射(约5%到达淋巴结)
●超声引导的淋巴结内注射
●静脉注射(很少能到达淋巴结)
DC疫苗剂量
0.3-3百万DC细胞是够的,但是比较精确的剂量还未知。
DC疫苗接种程序
肿瘤消退后,尽早接种以消除残留肿瘤灶。另外可以反复接种,以获得尽可能长的保护效果,是期望达到的效果。
DC疫苗的发展简史
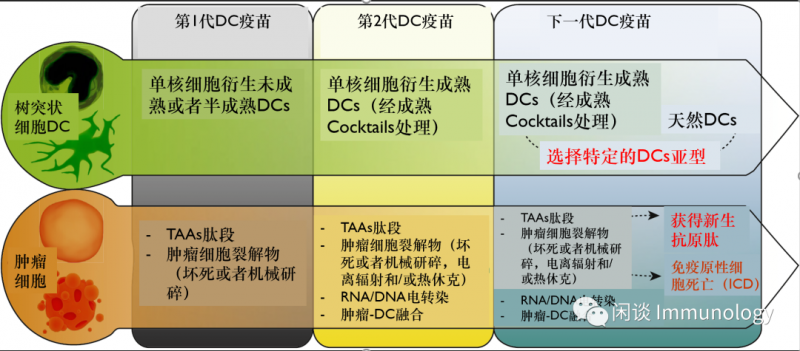
参考文献
[1] Steinman, R.M., Cohn, Z.A., 1973. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J. Exp. Med. 137,1142–1162.
[2] Schreibelt, G., 2012. The C-type lectin receptor CLEC9A mediates antigen uptake and (cross-)presentation by human blood BDCA3+ myeloid dendritic cells. Blood 119, 2284–2292.
[3] Cancel, J.C., Crozat, K., Dalod, M., 2019. Are conventional type 1 dendritic cells critical for protective antitumor immunity and how? Front. Immunol. 10, 9.
[4] Worah, K., 2016. Proteomics of human dendritic cell subsets reveals subset-specific surface markers and differential inflammasome function. Cell Rep. 16, 2953–2966.
[5] Swiecki, M., colonna, M., 2015. The multifaceted biology of plasmacytoid dendritic cells.Nat. Rev. Immunol. 15, 471–485.
[6] Reizis, B., 2019. Plasmacytoid dendritic cells: development, regulation, and function.Immunity 50, 37–50.
[7] Merad, M., Sathe, P., 2013. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 31, 563–604.
[8] Kubo, A., Nagao, K., 2009. External antigen uptake by Langerhans cells with reorganization of epidermal tight junction barriers. J. Exp. Med. 206, 2937–2946.
[9] Mansi Saxenaa, Nina Bhardwaja., 2019. Dendritic cell subsets and locations. International Review of Cell and Molecular Biology. 1937-6448.
[10] Salmon H, et al. Expansion and Activation of CD103(+) Dendritic Cell Progenitors at the Tumor Site Enhances Tumor Responses to Therapeutic PD-L1 and BRAF Inhibition. Immunity. 2016; 44(4):924–38.
[11] Thomas Naessens, et al. Human Lung Conventional Dendritic Cells Orchestrate Lymphoid Neogenesis During COPD. 2020; 2024(4).
[12] Garg, A. D., Coulie, P. G., Van den Eynde, B. J. & Agostinis, P. Integrating next- generation dendritic cell vaccines into the current cancer immunotherapy landscape. Trends Immunol. 38, 577–593 (2017).
[13] Stefanie K Wculek et al,Dendritic cells in cancer immunology and immunotherapy,Nat Rev Immunol . 2020 Jan;20(1):7-24.
[14] Kroemer, G. et al. (2013) Immunogenic cell death in cancer therapy. Ann. Rev. Immunol 31, 51–72
15 Aaes, T.L. et al. (2016) Vaccination with necroptotic cancer cells induces efficient anti-tumor immunity. Cell Rep. 15, 274–287
[16] Schumacher, T.N. and Schreiber, R.D. (2015) Neoantigens in cancer immunotherapy. Science 348, 69–74
[17] Kranz, L.M. et al. (2016) Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 534, 396–401
扫描进患者病友群

肺癌|胃癌|肝癌|结直肠癌|胰腺癌
乳腺癌|脑瘤|淋巴瘤|儿童肿瘤等

相关推荐
"癌王"胰腺癌不再无
EGFR突变型患者在非小细胞肺癌中约占30%,在欧美患者中约占10%~20%,在亚洲患者中占比超过50%,是非小细胞肺癌的第二大致癌驱动因素。...
全球肿瘤医生网 2020-08-05

多线耐药晚期肺癌有救了!MAGE-A10
EGFR突变型患者在非小细胞肺癌中约占30%,在欧美患者中约占10%~20%,在亚洲患者中占比超过50%,是非小细胞肺癌的第二大致癌驱动因素。...
全球肿瘤医生网 2020-08-05
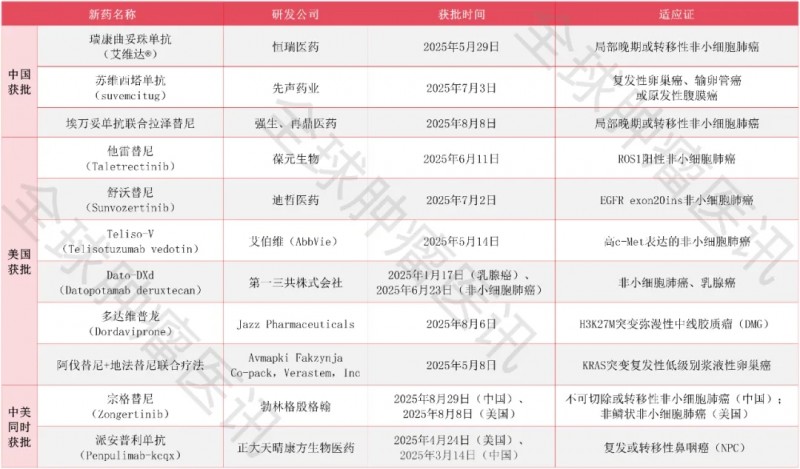
2025三季度抗癌"弹药库&q
EGFR突变型患者在非小细胞肺癌中约占30%,在欧美患者中约占10%~20%,在亚洲患者中占比超过50%,是非小细胞肺癌的第二大致癌驱动因素。...
全球肿瘤医生网 2020-08-05

愈见大"CAR"|C
EGFR突变型患者在非小细胞肺癌中约占30%,在欧美患者中约占10%~20%,在亚洲患者中占比超过50%,是非小细胞肺癌的第二大致癌驱动因素。...
全球肿瘤医生网 2020-08-05
全球首款!国产创新药舒沃替尼凭46%缓解
EGFR突变型患者在非小细胞肺癌中约占30%,在欧美患者中约占10%~20%,在亚洲患者中占比超过50%,是非小细胞肺癌的第二大致癌驱动因素。...
全球肿瘤医生网 2020-08-05

【KRAS药物】新一代靶向药HRS-70
EGFR突变型患者在非小细胞肺癌中约占30%,在欧美患者中约占10%~20%,在亚洲患者中占比超过50%,是非小细胞肺癌的第二大致癌驱动因素。...
全球肿瘤医生网 2020-08-05



![[field:title/]](/uploads/allimg/20200224/1-2002241I5262J.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241I3461D.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241H94LK.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241HKW34.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241H619327.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241H34H52.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241H22IH.jpg)
![[field:title/]](/uploads/allimg/20200224/1-2002241G93O18.jpg)




